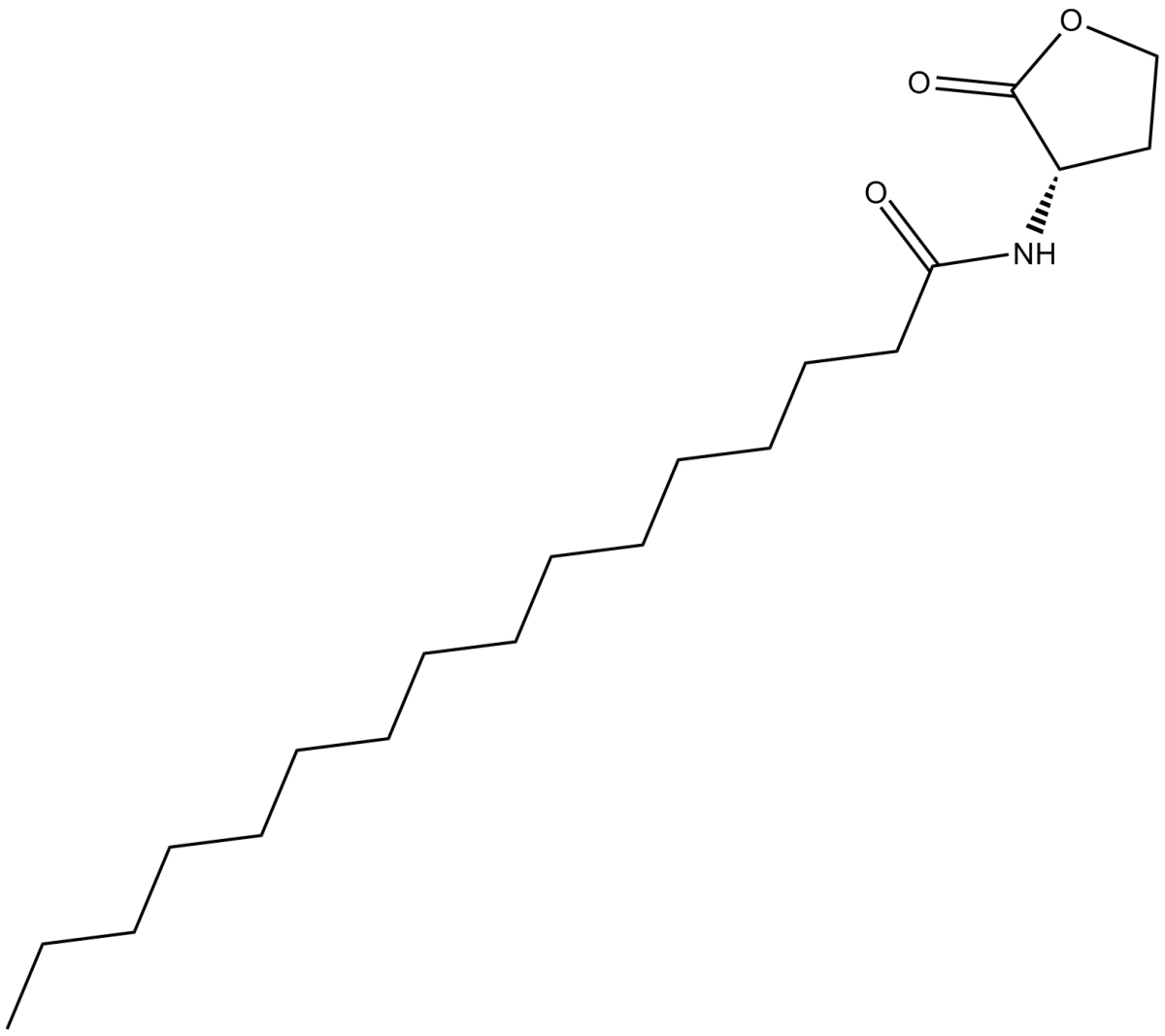
| 产品描述 | Quorum sensing is a regulatory system used by bacteria for controlling gene expression in response to increasing cell density.[1] This regulatory process manifests itself with a variety of phenotypes including biofilm formation and virulence factor production.[2] Coordinated gene expression is achieved by the production, release, and detection of small diffusible signal molecules called autoinducers. The N-acylated homoserine lactones (AHLs) comprise one such class of autoinducers, each of which generally consists of a fatty acid coupled with homoserine lactone (HSL). Regulation of bacterial quorum sensing signaling systems to inhibit pathogenesis represents a new approach to antimicrobial therapy in the treatment of infectious diseases.[3] AHLs vary in acyl group length (C4-C18), in the substitution of C3 (hydrogen, hydroxyl, or oxo group), and in the presence or absence of one or more carbon-carbon double bonds in the fatty acid chain. These differences confer signal specificity through the affinity of transcriptional regulators of the LuxR family.[4] C16-HSL is one of a number of lipophilic, long acyl side-chain bearing AHLs, including its monounsaturated analog C16:1-(L)-HSL, produced by the LuxI AHL synthase homolog SinI involved in quorum-sensing signaling in S. meliloti, a nitrogen-fixing bacterial symbiont of certain legumes.[5],[6] C16-HSL is the most abundant AHL produced by the proteobacterium R. capsulatus and activates genetic exchange between R. capsulatus cells.[7] N-Hexadecanoyl-L-homoserine lactone and other hydrophobic AHLs tend to localize in relatively lipophilic cellular environments of bacteria and cannot diffuse freely through the cell membrane. The long-chain N-acylhomoserine lactones may be exported from cells by efflux pumps or may be transported between communicating cells by way of extracellular outer membrane vesicles.[8],[9]
Reference:
[1]. González, J.E., and Keshavan, N.D. Messing with bacterial quorum sensing Microbiol. Mol. Biol. Rev. 70(4), 859-875 (2006).
[2]. Gould, T.A., Herman, J., Krank, J., et al. Specificity of acyl-homoserine lactone syntheses examined by mass spectrometry Journal of Bacteriology 188(2), 773-783 (2006).
[3]. Cegelski, L., Marshall, G.R., Eldridge, G.R., et al. The biology and future prospects of antivirulence therapies Nature Reviews.Microbiology 6(1), 17-27 (2008).
[4]. Penalver, C.G.N., Morin, D., Cantet, F., et al. Methylobacterium extorquens AM1 produces a novel type of acyl-homoserine lactone with a double unsaturated side chain under methylotrophic growth conditions FEBS Letters 580, 561-567 (2006).
[5]. Gao, M., Chen, H., Eberhard, A., et al. sinI- and expR-dependent quorum sensing in Sinorhizobium meliloti Journal of Bacteriology 187(23), 7931-7944 (2005).
[6]. Teplitski, M., Eberhard, A., Gronquist, M.R., et al. Chemical identification of N-acyl homoserine lactone quorum-sensing signals produced by Sinorhizobium meliloti strains in defined medium Archives of Microbiology 180, 494-497 (2003).
[7]. Schaefer, A.L., Taylor, T.A., Beatty, J.T., et al. Long-chain acyl-homoserine lactone quorum-sensing regulation of Rhodobacter capsulatus gene transfer agent production Journal of Bacteriology 184(23), 6515-6521 (2002).
[8]. Pearson, J.P., Van Delden, C., and Iglewski, B.H. Active efflux and diffusion are involved in transport of Pseudomonas aeruginosa cell-to-cell signals Journal of Bacteriology 181(4), 1203-1210 (1999).
[9]. Mashburn-Warren, L., and Whiteley, M. Special delivery: Vesicle trafficking in prokaryotes Molecular Microbiology 61(4), 839-846 (2006). |
 m.cnreagent.com
m.cnreagent.com